
Весь контент iLive перевіряється медичними експертами, щоб забезпечити максимально можливу точність і відповідність фактам.
У нас є строгі правила щодо вибору джерел інформації та ми посилаємося тільки на авторитетні сайти, академічні дослідницькі інститути і, по можливості, доведені медичні дослідження. Зверніть увагу, що цифри в дужках ([1], [2] і т. д.) є інтерактивними посиланнями на такі дослідження.
Якщо ви вважаєте, що який-небудь з наших матеріалів є неточним, застарілим або іншим чином сумнівним, виберіть його і натисніть Ctrl + Enter.
Гіпофіз
Медичний експерт статті

Останній перегляд: 04.07.2025

Гіпофіз (hypophysis, s.glandula pituitaria) розташований у гіпофізарній ямці турецького сідла клиноподібної кістки та відокремлений від порожнини черепа відростком твердої мозкової оболонки, утворюючи діафрагму сідла. Через отвір у цій діафрагмі гіпофіз з'єднується з воронкою гіпоталамуса проміжного мозку. Поперечний розмір гіпофіза становить 10-17 мм, передньозадній - 5-15 мм, вертикальний - 5-10 мм. Маса гіпофіза у чоловіків становить приблизно 0,5 г, у жінок - 0,6 г. Зовні гіпофіз покритий капсулою.
Відповідно до розвитку гіпофіза з двох різних зачатків, в органі розрізняють дві частки - передню та задню. Аденогіпофіз, або передня частка (adenohypophysis, s.lobus anterior), більша, становить 70-80% від загальної маси гіпофіза. Вона щільніша за задню частку. У передній частці розрізняють дистальну частину (pars distalis), яка займає передню частину гіпофізарної ямки, проміжну частину (pars intermedia), розташовану на межі із задньою часткою, та горбисту частину (pars tuberalis), що йде вгору та з'єднується з воронкою гіпоталамуса. Завдяки великій кількості кровоносних судин передня частка має блідо-жовтий колір з червонуватим відтінком. Паренхіма передньої частини гіпофіза представлена кількома типами залозистих клітин, між тяжами яких розташовані синусоїдальні кровоносні капіляри. Половина (50%) клітин аденогіпофіза – це хромофільні аденоцити, які мають дрібнозернисті гранули в цитоплазмі, що добре забарвлюються солями хрому. Це ацидофільні аденоцити (40% усіх клітин аденогіпофіза) та базофільні аденоцити (10%). До базофільних аденоцитів належать гонадотропні, кортикотропні та тиреотропні ендокриноцити. Хромофобні аденоцити мають малі розміри, велике ядро та невелику кількість цитоплазми. Ці клітини вважаються попередниками хромофільних аденоцитів. Інші 50% клітин аденогіпофіза – це хромофобні аденоцити.
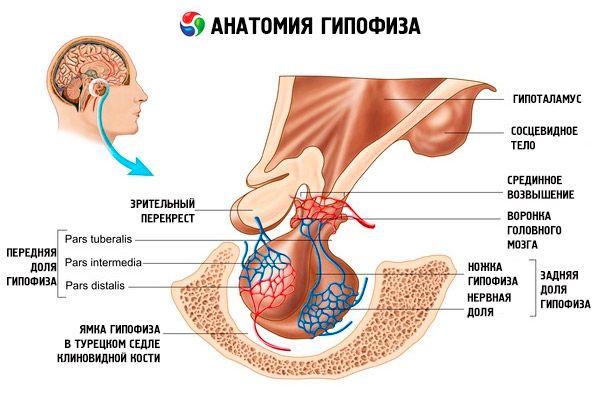
Нейрогіпофіз, або задня частка (neurohypophysis, s.lobus posterior), складається з нервової частки (lobus nervosus), яка розташована в задній частині гіпофізарної ямки, та воронки (infundibulum), розташованої позаду горбистої частини аденогіпофіза. Задня частка гіпофіза утворена нейрогліальними клітинами (клітинами гіпофіза), нервовими волокнами, що йдуть від нейросекреторних ядер гіпоталамуса до нейрогіпофіза, та нейросекреторними тільцями.
Гіпофіз, через нервові волокна (шляхи) та кровоносні судини, функціонально пов'язаний з гіпоталамусом проміжного мозку, який регулює діяльність гіпофіза. Гіпофіз та гіпоталамус разом з їхніми нейроендокринними, судинними та нервовими зв'язками зазвичай вважаються гіпоталамо-гіпофізарною системою.
Гормони передньої та задньої долі гіпофіза впливають на багато функцій організму, насамперед через інші ендокринні залози. У передній долі гіпофіза ацидофільні аденоцити (альфа-клітини) виробляють соматотропний гормон (гормон росту), який бере участь у регуляції процесів росту та розвитку у молодих організмів. Кортикотропні ендокриноцити секретують адренокортикотропний гормон (АКТГ), який стимулює секрецію стероїдних гормонів наднирковими залозами. Тиреотропні ендокриноцити секретують тиреотропний гормон (ТТГ), який впливає на розвиток щитовидної залози та активує вироблення її гормонів. Гонадотропні гормони: фолікулостимулюючий (ФСГ), лютеїнізуючий (ЛГ) та пролактин – впливають на статеве дозрівання організму, регулюють та стимулюють розвиток фолікулів у яєчнику, овуляцію, ріст молочних залоз та вироблення молока у жінок, процес сперматогенезу у чоловіків. Ці гормони виробляються базофільними аденоцитами (бета-клітинами). Тут також секретуються ліпотропні фактори гіпофіза, які впливають на мобілізацію та утилізацію жирів в організмі. У проміжній частині передньої частки утворюється меланоцитстимулюючий гормон, який контролює утворення пігментів – меланінів – в організмі.
Нейросекреторні клітини супраоптичного та паравентрикулярного ядер у гіпоталамусі виробляють вазопресин та окситоцин. Ці гормони транспортуються до клітин задньої частки гіпофіза по аксонах, що утворюють гіпоталамо-гіпофізний тракт. Із задньої частки гіпофіза ці речовини потрапляють у кров. Гормон вазопресин має вазоконстрикторну та антидіуретичну дію, за що його також називають антидіуретичним гормоном (АДГ). Окситоцин має стимулюючий вплив на скоротливість м'язів матки, збільшує секрецію молока лактуючою молочною залозою, пригнічує розвиток та функцію жовтого тіла, впливає на зміни тонусу гладкої (несмугастої) мускулатури шлунково-кишкового тракту.
Розвиток гіпофіза
Передня частка гіпофіза розвивається з епітелію дорсальної стінки ротової затоки у вигляді кільцеподібного виросту (кишеня Ратке). Це ектодермальне випинання зростає до дна майбутнього третього шлуночка. До нього, від нижньої поверхні другого мозкового міхура (майбутнього дна третього шлуночка), зростає відросток, з якого розвиваються сірий горбок воронки та задня частка гіпофіза.
Судини та нерви гіпофіза
Від внутрішніх сонних артерій та судин артеріального кола великого мозку верхня та нижня гіпофізарні артерії направляються до гіпофіза. Верхні гіпофізарні артерії йдуть до сірого ядра та воронки гіпоталамуса, анастомозують тут одна з одною та утворюють капіляри, що проникають у тканину мозку – первинну гемокапілярну мережу. З довгих та коротких петель цієї мережі утворюються ворітні вени, які направляються до передньої частки гіпофіза. У паренхімі передньої частки гіпофіза ці вени розпадаються на широкі синусоїдальні капіляри, утворюючи вторинну гемокапілярну мережу. Задня частка гіпофіза кровопостачається переважно нижньою гіпофізарною артерією. Між верхньою та нижньою гіпофізарними артеріями є довгі артеріальні анастомози. Відтік венозної крові з вторинної гемокапілярної мережі здійснюється через систему вен, що впадають у печеристі та міжпещирні синуси твердої мозкової оболонки мозку.
У іннервації гіпофіза беруть участь симпатичні волокна, що входять в орган разом з артеріями. Постгангліонарні симпатичні нервові волокна відходять від сплетення внутрішньої сонної артерії. Крім того, в задній частці гіпофіза знаходяться численні закінчення відростків нейросекреторних клітин, розташованих у ядрах гіпоталамуса.
Вікові особливості гіпофіза
Середня вага гіпофіза у новонароджених сягає 0,12 г. Вага органу подвоюється до 10 років і потроюється до 15 років. До 20 років вага гіпофіза досягає свого максимуму (530-560 мг) і залишається майже незмінною в наступні вікові періоди. Після 60 років спостерігається незначне зменшення ваги цієї ендокринної залози.
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
Гормони гіпофіза
Єдність нервової та гормональної регуляції в організмі забезпечується тісним анатомічно-функціональним зв'язком гіпофіза та гіпоталамуса. Цей комплекс визначає стан і функціонування всієї ендокринної системи.
Основною ендокринною залозою, що виробляє ряд пептидних гормонів, що безпосередньо регулюють функцію периферичних залоз, є гіпофіз. Це червонувато-сіре бобоподібне утворення, вкрите фіброзною капсулою вагою 0,5-0,6 г. Воно дещо змінюється залежно від статі та віку людини. Залишається загальноприйнятим поділ гіпофіза на дві частки, що відрізняються розвитком, будовою та функцією: передню дистальну - аденогіпофіз та задню - нейрогіпофіз. Перша становить близько 70% від загальної маси залози та умовно поділяється на дистальну, воронкоподібну та проміжну частини, друга - на задню частину, або частку, та ніжку гіпофіза. Залоза розташована в гіпофізарній ямці турецького сідла клиноподібної кістки та з'єднана з мозком через ніжку. Верхня частина передньої частки покрита зоровим перехрестям та зоровими трактами. Кровопостачання гіпофіза дуже рясне і здійснюється гілками внутрішньої сонної артерії (верхня та нижня гіпофізарні артерії), а також гілками артеріального кола великого мозку. Верхні гіпофізарні артерії беруть участь у кровопостачанні аденогіпофіза, а нижні - нейрогіпофіза, контактуючи з нейросекреторними закінченнями аксонів великоклітинних ядер гіпоталамуса. Перші входять у серединне піднесення гіпоталамуса, де розходяться в капілярну мережу (первинне капілярне сплетення). Ці капіляри (з якими контактують закінчення аксонів дрібних нейросекреторних клітин медіобазального гіпоталамуса) збираються в ворітні вени, що спускаються вздовж ніжки гіпофіза в паренхіму аденогіпофіза, де знову поділяються на мережу синусоїдальних капілярів (вторинне капілярне сплетення). Таким чином, кров, попередньо пройшовши через серединне піднесення гіпоталамуса, де вона збагачується гіпоталамічними аденогіпофізотропними гормонами (рилізинг-гормонами), потрапляє в аденогіпофіз.
Відтік крові, насиченої гормонами аденогіпофіза, з численних капілярів вторинного сплетення здійснюється через систему вен, які в свою чергу впадають у венозні синуси твердої мозкової оболонки, а потім у загальний кровотік. Таким чином, портальна система гіпофіза з низхідним напрямком кровотоку від гіпоталамуса є морфофункціональним компонентом складного механізму нейрогуморального контролю тропних функцій аденогіпофіза.
Гіпофіз іннервується симпатичними волокнами, що йдуть вздовж гіпофізарних артерій. Вони беруть початок від постгангліонарних волокон, що проходять через внутрішнє сонне сплетення, з'єднане з верхніми шийними гангліями. Безпосередньої іннервації аденогіпофіза з гіпоталамуса немає. Задня частка отримує нервові волокна від нейросекреторних ядер гіпоталамуса.
Аденогіпофіз — це дуже складне за своєю гістологічною архітектурою утворення. Він має два типи залозистих клітин — хромофобні та хромофільні. Останні, у свою чергу, поділяються на ацидофільні та базофільні (детальний гістологічний опис гіпофіза наведено у відповідному розділі посібника). Однак слід зазначити, що гормони, що виробляються залозистими клітинами, що складають паренхіму аденогіпофіза, через різноманітність останніх певною мірою відрізняються за своєю хімічною природою, а тонка структура секретуючих клітин повинна відповідати особливостям біосинтезу кожної з них. Але іноді в аденогіпофізі можна спостерігати перехідні форми залозистих клітин, які здатні виробляти кілька гормонів. Є дані, що тип залозистих клітин аденогіпофіза не завжди визначається генетично.
Під діафрагмою турецького сідла розташована лійкоподібна частина передньої частки. Вона охоплює ніжку гіпофіза, контактуючи з сірим горбком. Ця частина аденогіпофіза характеризується наявністю епітеліальних клітин та рясним кровопостачанням. Вона також гормонально активна.
Проміжна (середня) частина гіпофіза складається з кількох шарів великих секреторно-активних базофільних клітин.
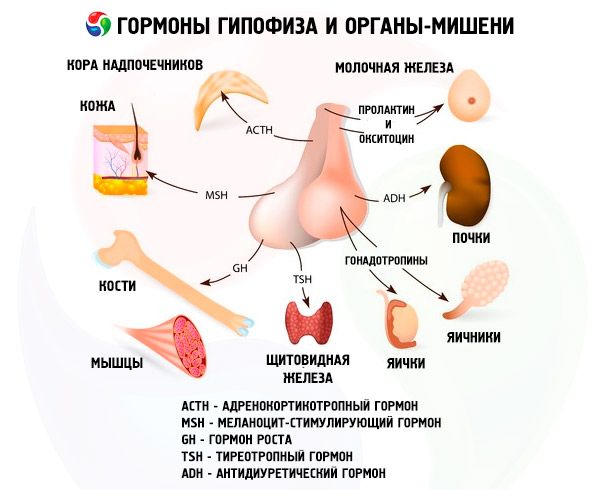
Гіпофіз виконує різні функції за допомогою своїх гормонів. Його передня частка виробляє адренокортикотропний (АКТГ), тиреостимулюючий (ТТГ), фолікулостимулюючий (ФСГ), лютеїнізуючий (ЛГ), ліпотропні гормони, а також гормон росту – соматотропний (СТО) та пролактин. У проміжній частці синтезується меланоцитстимулюючий гормон (МСГ), а в задній частці накопичуються вазопресин та окситоцин.
АКТГ
Гормони гіпофіза – це група білкових і пептидних гормонів і глікопротеїнів. З гормонів передньої долі гіпофіза найкраще вивчений АКТГ. Він виробляється базофільними клітинами. Його основна фізіологічна функція полягає у стимуляції біосинтезу та секреції стероїдних гормонів корою надниркових залоз. АКТГ також проявляє меланоцитостимулюючу та ліпотропну активність. У 1953 році його було виділено в чистому вигляді. Пізніше було встановлено його хімічну структуру, що складається з 39 амінокислотних залишків у людини та ряду ссавців. АКТГ не має видової специфічності. Наразі проведено хімічний синтез як самого гормону, так і різних фрагментів його молекули, більш активних, ніж природні гормони. Структура гормону має дві ділянки пептидного ланцюга, одна з яких забезпечує виявлення та зв'язування АКТГ з рецептором, а інша забезпечує біологічний ефект. Він, очевидно, зв'язується з рецептором АКТГ шляхом взаємодії електричних зарядів гормону та рецептора. Роль біологічного ефектора АКТГ виконує фрагмент молекули 4-10 (Met-Glu-His-Phen-Arg-Tri-Tri).
Меланоцитстимулююча активність АКТГ зумовлена наявністю в молекулі N-кінцевої ділянки, що складається з 13 амінокислотних залишків і повторює структуру альфа-меланоцитстимулюючого гормону. Ця ділянка також містить гептапептид, присутній в інших гормонах гіпофіза та володіє деякою адренокортикотропною, меланоцитстимулюючою та ліпотропною активністю.
Ключовим моментом у дії АКТГ вважається активація ферменту протеїнкінази в цитоплазмі за участю цАМФ. Фосфорильована протеїнкіназа активує фермент естеразу, яка перетворює ефіри холестерину на вільну речовину в жирових краплях. Білок, синтезований у цитоплазмі в результаті фосфорилювання рибосом, стимулює зв'язування вільного холестерину з цитохромом Р-450 та його перенесення з ліпідних крапель до мітохондрій, де присутні всі ферменти, що забезпечують перетворення холестерину на кортикостероїди.
[ 16 ], [ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ], [ 22 ]
Тиреотропний гормон
ТТГ – тиреотропін – основний регулятор розвитку та функціонування щитовидної залози, процесів синтезу та секреції гормонів щитовидної залози. Цей складний білок – глікопротеїн – складається з альфа- та бета-субодиниць. Структура першої субодиниці збігається з альфа-субодиницею лютеїнізуючого гормону. Більше того, вона значною мірою збігається у різних видів тварин. Послідовність амінокислотних залишків у бета-субодиниці ТТГ людини розшифрована і складається зі 119 амінокислотних залишків. Можна зазначити, що бета-субодиниці ТТГ людини та великої рогатої худоби багато в чому схожі. Біологічні властивості та характер біологічної активності глікопротеїнових гормонів визначаються бета-субодиницею. Вона також забезпечує взаємодію гормону з рецепторами в різних органах-мішенях. Однак бета-субодиниця у більшості тварин проявляє специфічну активність лише після з'єднання з альфа-субодиницею, яка виступає своєрідним активатором гормону. Остання з однаковою ймовірністю індукує лютеїнізуючу, фолікулостимулюючу та тиреотропну активності, що визначаються властивостями бета-субодиниці. Виявлена подібність дозволяє зробити висновок, що ці гормони виникли в процесі еволюції від одного спільного попередника, бета-субодиниця також визначає імунологічні властивості гормонів. Існує припущення, що альфа-субодиниця захищає бета-субодиницю від дії протеолітичних ферментів, а також сприяє її транспортуванню з гіпофіза до периферичних органів-мішеней.
Гонадотропні гормони
Гонадотропіни присутні в організмі у формі ЛГ та ФСГ. Функціональне призначення цих гормонів загалом зводиться до забезпечення репродуктивних процесів у особин обох статей. Вони, як і ТТГ, є складними білками - глікопротеїнами. ФСГ індукує дозрівання фолікулів у яєчниках жінок та стимулює сперматогенез у чоловіків. ЛГ викликає розрив фолікулів у жінок з утворенням жовтого тіла та стимулює секрецію естрогенів та прогестерону. У чоловіків цей самий гормон прискорює розвиток інтерстиціальної тканини та секрецію андрогенів. Вплив гонадотропінів залежить один від одного та відбувається синхронно.
Динаміка секреції гонадотропінів у жінок змінюється протягом менструального циклу та вивчена достатньо детально. У преовуляторній (фолікулярній) фазі циклу вміст ЛГ знаходиться на досить низькому рівні, тоді як ФСГ підвищений. У міру дозрівання фолікула секреція естрадіолу збільшується, що сприяє підвищенню вироблення гонадотропінів гіпофізом та виникненню як циклів ЛГ, так і ФСГ, тобто статеві стероїди стимулюють секрецію гонадотропінів.
Наразі структура ЛГ визначена. Як і ТТГ, він складається з 2 субодиниць: a та b. Структура альфа-субодиниці ЛГ у різних видів тварин значною мірою збігається, вона відповідає структурі альфа-субодиниці ТТГ.
Структура бета-субодиниці ЛГ суттєво відрізняється від структури бета-субодиниці ТТГ, хоча вона має чотири однакові ділянки пептидного ланцюга, що складаються з 4-5 амінокислотних залишків. У ТТГ вони локалізовані в положеннях 27-31, 51-54, 65-68 та 78-83. Оскільки бета-субодиниця ЛГ та ТТГ визначає специфічну біологічну активність гормонів, можна припустити, що гомологічні ділянки в структурі ЛГ та ТТГ повинні забезпечувати зв'язок бета-субодиниць з альфа-субодиницею, а ділянки, що відрізняються за структурою, повинні відповідати за специфічність біологічної активності гормонів.
Нативний ЛГ дуже стійкий до дії протеолітичних ферментів, але бета-субодиниця швидко розщеплюється хімотрипсином, а α-субодиниця важко гідролізується ферментом, тобто вона відіграє захисну роль, запобігаючи доступу хімотрипсину до пептидних зв'язків.
Що стосується хімічної структури ФСГ, дослідники ще не отримали остаточних результатів. Як і ЛГ, ФСГ складається з двох субодиниць, але бета-субодиниця ФСГ відрізняється від бета-субодиниці ЛГ.
Пролактин
Інший гормон, пролактин (лактогенний гормон), відіграє активну роль у процесах розмноження. Основні фізіологічні властивості пролактину у ссавців проявляються у вигляді стимуляції розвитку молочних залоз та лактації, росту сальних залоз та внутрішніх органів. Він сприяє прояву впливу стероїдів на вторинні статеві ознаки у самців, стимулює секреторну активність жовтого тіла у мишей та щурів, бере участь у регуляції жирового обміну. В останні роки пролактину приділяється велика увага як регулятору материнської поведінки; така поліфункціональність пояснюється його еволюційним розвитком. Він є одним із найдавніших гормонів гіпофіза і зустрічається навіть у земноводних. Наразі структура пролактину у деяких видів ссавців повністю розшифрована. Однак донедавна вчені висловлювали сумніви щодо існування такого гормону у людини. Багато хто вважав, що його функцію виконує гормон росту. Зараз отримано переконливі докази наявності пролактину у людини, і його структура частково розшифрована. Рецептори пролактину активно зв'язуються з гормоном росту та плацентарним лактогеном, що вказує на єдиний механізм дії трьох гормонів.
Соматотропін
Гормон росту, соматотропін, має ще ширший спектр дії, ніж пролактин. Як і пролактин, він виробляється ацидофільними клітинами аденогіпофіза. СТГ стимулює ріст скелета, активує біосинтез білка, має жиромобілізуючу дію та сприяє збільшенню розмірів тіла. Крім того, він координує метаболічні процеси.
Участь гормону в останньому підтверджується фактом різкого збільшення його секреції гіпофізом, наприклад, при зниженні рівня цукру в крові.
Хімічна структура цього людського гормону зараз повністю встановлена — 191 амінокислотний залишок. Його первинна структура подібна до структури хоріонічного соматомаммотропіну або плацентарного лактогену. Ці дані свідчать про значну еволюційну близькість двох гормонів, хоча вони демонструють відмінності в біологічній активності.
Необхідно підкреслити високу видову специфічність розглянутого гормону – наприклад, СТГ тваринного походження неактивний у людини. Це пояснюється як реакцією між рецепторами СТГ людини та тварин, так і структурою самого гормону. Наразі проводяться дослідження з виявлення активних центрів у складній структурі СТГ, що проявляють біологічну активність. Вивчаються окремі фрагменти молекули, що проявляють інші властивості. Наприклад, після гідролізу людського СТГ пепсином було виділено пептид, що складається з 14 амінокислотних залишків і відповідає ділянці молекули 31-44. Він не мав ростового ефекту, але значно перевершував нативний гормон за ліпотропною активністю. Гормон росту людини, на відміну від аналогічного гормону тварин, має значну лактогенну активність.
Аденогіпофіз синтезує багато пептидних і білкових речовин, що мають жиромобілізуючу дію, а ліпотропний ефект мають тропні гормони гіпофіза – АКТГ, СТГ, ТТГ та інші. В останні роки особливо підкреслюються бета- та γ-ліпотропні гормони (ЛПГ). Найбільш детально вивчені біологічні властивості бета-ЛПГ; крім ліпотропної активності, він також має меланоцитстимулюючу, кортикотропінстимулюючу та гіпокальціємічну дію, а також виробляє інсуліноподібний ефект.
Наразі розшифровано первинну структуру LPG овець (90 амінокислотних залишків), ліпотропних гормонів свиней та великої рогатої худоби. Цей гормон має видову специфічність, хоча структура центральної області бета-LPG однакова у різних видів. Це визначає біологічні властивості гормону. Один з фрагментів цієї області виявлено в структурі альфа-MSH, бета-MSH, АКТГ та бета-LPG. Припускають, що ці гормони виникли з одного попередника в процесі еволюції. γ-LPG має слабшу ліпотропну активність, ніж бета-LPG.
Меланоцитстимулюючий гормон
Цей гормон, що синтезується в проміжній частці гіпофіза, стимулює біосинтез шкірного пігменту меланіну в його біологічній функції, сприяє збільшенню розміру та кількості пігментних клітин меланоцитів у шкірі амфібій. Ці якості МСГ використовуються в біологічних випробуваннях гормону. Існує два типи гормону: альфа- та бета-МСГ. Було показано, що альфа-МСГ не має видової специфічності та має однакову хімічну структуру у всіх ссавців. Його молекула являє собою пептидний ланцюг, що складається з 13 амінокислотних залишків. Бета-МСГ, навпаки, має видову специфічність, і його структура відрізняється у різних тварин. У більшості ссавців молекула бета-МСГ складається з 18 амінокислотних залишків, і лише у людини вона подовжена від амінокислотного кінця на чотири амінокислотні залишки. Слід зазначити, що альфа-МСГ має певну адренокортикотропну активність, і його вплив на поведінку тварин і людини зараз доведено.
Окситоцин і вазопресин
Вазопресин та окситоцин, що синтезуються в гіпоталамусі, накопичуються в задній частці гіпофіза: вазопресин у нейронах супраоптичного ядра, а окситоцин у паравентрикуляторному ядрі. Потім вони переносяться до гіпофіза. Слід підкреслити, що попередник гормону вазопресину синтезується спочатку в гіпоталамусі. При цьому там виробляються білки нейрофізини 1 та 2 типів. Перший зв'язує окситоцин, а другий - вазопресин. Ці комплекси мігрують у вигляді нейросекреторних гранул у цитоплазмі вздовж аксона та досягають задньої частки гіпофіза, де нервові волокна закінчуються в судинній стінці, а вміст гранул потрапляє в кров. Вазопресин та окситоцин - перші гормони гіпофіза з повністю встановленою амінокислотною послідовністю. За своєю хімічною структурою вони є нонапептидами з одним дисульфідним містком.
Розглянуті гормони мають різні біологічні ефекти: вони стимулюють транспорт води та солей через мембрани, мають вазопресорну дію, посилюють скорочення гладкої мускулатури матки під час пологів та збільшують секрецію молочних залоз. Слід зазначити, що вазопресин має вищу антидіуретичну активність, ніж окситоцин, тоді як останній має сильніший вплив на матку та молочну залозу. Основним регулятором секреції вазопресину є споживання води; у ниркових канальцях він зв'язується з рецепторами в цитоплазматичних мембранах з подальшою активацією в них ферменту аденілатциклази. За зв'язування гормону з рецептором та за біологічний ефект відповідають різні частини молекули.
Гіпофіз, пов'язаний через гіпоталамус з усією нервовою системою, об'єднує ендокринну систему в функціональне ціле, беручи участь у забезпеченні сталості внутрішнього середовища організму (гомеостазу). У межах ендокринної системи гомеостатична регуляція здійснюється на основі принципу зворотного зв'язку між передньою часткою гіпофіза та залозами-"мішенями" (щитовидною залозою, корою надниркових залоз, статевими залозами). Надлишок гормону, що виробляється залозою-"мішенями", пригнічує, а його дефіцит стимулює секрецію та виділення відповідного тропного гормону. Гіпоталамус включений у систему зворотного зв'язку. Саме в ньому розташовані рецепторні зони, чутливі до гормонів залоз-"мішеней". Специфічно зв'язуючись з гормонами, що циркулюють у крові, та змінюючи реакцію залежно від концентрації гормонів, гіпоталамічні рецептори передають свій вплив відповідним гіпоталамічним центрам, які координують роботу аденогіпофіза, вивільняючи гіпоталамічні аденогіпофізіотропні гормони. Таким чином, гіпоталамус слід розглядати як нейроендокринний мозок.
Использованная литература